Concept of maintenance breeding in cotton
Maintenance breeding is a branch of breeding, implemented to maintain the genetic purity, vigor, yield potential and adoptation of a variety or a parent. There are numerous examples of varieties lost their yield potential due to the lack of maintenance breeding (Peng. 2010). The importance of maintenance breeding can be easily understood by investigating the genetic cause behind the loss of yield potential.
Cotton is the often cross pollinated crop with predominance of self pollination and low amount of (~5% ) cross pollination. This type of pollination system has two important features. In open pollination, there will be a free exchange of pollens between plants. Secondly, Expression of heterosis (increase of hybrid vigor due to the heterozygosity of the Hybrid). The different alleles of the same gene contributes more to the phenotype at heterozygous state than at homozygous due to over dominance of alleles. The genes at homozygous state leads to inbreeding depression or loss of hybrid vigor.
The genetics of an open pollinating pureline
A variety is a ‘pure line’. The progenies of a pureline are homozygous and homogenous in nature. Seed multiplication in cotton is taken at isolation fields and maintained by open pollination. Though this population is said to be homozygous and homogenous, this is not the actual case.
The genetic makeup of an open pollinating variety will be
- Homozygous for morphological/qualitative traits
- Heterozygous for agronomical/Quantitative traits
The qualitative characters are controlled by one or few genes, and quantitative traits are controlled by polygene. The varietal development breeding involves crossing of two parents followed by subsequent generations of selection and selfing till the selected line attains homozygosity. The homozygous progeny is now a pureline. The homozygosity of qualitative genes leads to uniformity in the morphological traits. At the same time the agronomical traits suffers inbreeding depression due to the homozygosity in polygenic alleles.
When the variety is brought to the open pollination from selfing, the variety regains its vigor in one cycle of open pollination. This is evident from the increased vigor of the progenies of the open pollinated variety. This clearly indicates the heterozygosity in polygenic traits are important for the maintenance of ‘varietal heterosis’ for vigor and yield potential.Out crossing in cotton is reported occur at the proportion of ~5%. Most of the out crossings occur within one meter of distance. Hence each plant receives ~5% pollen from the nearby plants leads to the heterozygosity of quantitative loci.
The Genotype x environment (G x E) is the another important characteristic of polygenic traits. The environment influences more than 30% of the phenotypic expression of polygenes. Progenies of a pureline will differ in the G x E interaction ability. To put it in simple, consider, if two plants has same genotype, grown under same environment, the two plants might not have the same interaction with the environment, and the ‘fittest’ plants potential will be little more than the other plant.
What happens when there is no ‘Maintenance Breeding’?
Loss of genetic purity
Genetic purity is uniformity of plants in terms of morphological traits. Any variation in the qualitative traits indicates disturbance of genetic purity. In my previous blog, I have explained the appearance of such ‘off type’ plants in the seed multiplication plot. Heterozygosity, outcrossing, and point mutation are agents, creating variation in a pure line. If the variations are un noticed over the generations, result in the amplification of off type plants, and loss of genetic purity
Loss of vigor and potential
As we mentioned earlier, heterozygosity at loci of agronomical traits are important for maintaining the vigor. Under continuous selfing, the variety suffers inbreeding depression. The open pollination restores the heterosis or vigor of that variety due to some extend of outcrossing with nearby plants. But outcrossing is very limited in cotton and self-pollination is predominant. This amount of pollen exchange is not sufficient to get desired heterozygosity to retain the yield potential of the variety. Without maintenance breeding, the variety will approach the homozygous state even under open pollination, causes inbreeding depression
Loss of fitness/adaptation
Genotype x Environmental interactions is the measure of a genotypes ability to adopt for the changing environmental conditions. The environmental conditions changes over the seasons. The temperature, drought and salinity increases every year, and it has negative impact on a genotypes vigor and potential. If the variety in not maintained under changing environments, the relative fitness of the variety will get drastically reduced.
How maintenance breeding is done
The maintenance of a variety requires maintenance of genetic purity, vigor and fitness separately but altogether in the same plot.
Maintaining genetic purity:
The visible deviations in the morphological traits can be easily identified by the careful inspection of the plot. The off type plants should be immediately removed from the field to avoid the out crossing with nearby ‘pure’ plants. If the off type plants proportion is less than 0.01%, the removal of those plants is enough to maintain the genetic purity. Because the off type plants might be the results of either natural out crossing or by mutation. If the off type proportion is more than 0.1%, indicates the heterozygosity of few plants in the source seeds. In this case removal of off type plants are not enough, since some plants will be at heterozygous and escapes the visible detection of off phenotypes. In such situations, the genetic purity can be maintained by selection of pure plants form the plot and selfing of the plants. The selfed seeds are raised in the next season in plant to row method. The plants of a segregating progenies are removed from the plot and only the non-segregating uniform progenies are retained and allowed for open pollination.
Maintenance of vigor and yield potential
The inbreeding depression due to the preponderance of self-pollination can be alleviated by sib mating of individual plants. The best performing plants are selected and crossing is performed between all selected plants in all combinations. Each plant is used as a male as well as female to restore the cytoplasm of the all selected plants. Sib crossing ensures the union of alleles of different plants. The progenies of sib mated plants expresses the heterosis in vigor due to the higher level of heterozygosity in the quantitative loci
Maintenance of fitness
To maintain the fitness of a variety, it should be routinely brought in to the environment and perfoamce is monitored. The variety maintained by proper roguing and sib mating, might have high genetic purity and uniform vigor but differs in fitness levels. productivity of some plants are superior to the other plants due to positive G x E interactions. Selection of superior plants increases the adoptability of the genotypes to that particular environment. The important point to note here is, selection will not improve the superiority of the genotypes. (“Selection is not effective within a pure line”. Johansen. 1906). Selection is only effective for the retention of ‘superiority’. When the best performing plants are selected under particular environmental conditions, those plants will express more fitness in the successive generations of varietal Maintenance. To increase the longevity of the fitness, the variety should be maintained once in two years under the particular environmental conditions and best interacting (G x E) plants should be selected as the source for next round of multiplication.
Procedure for Maintenance Breeding
One cycle of maintenance breeding involves
Year 1
Raising the variety in isolation fields under open pollination, rouging of off type plants, selection of best plants and sib mating between the selected plants
Year 2
Raising the sib progenies, rouging of off type plants, and selection of best plants.
After one or two years, the cycle should be started again. The selected plants at the end of year 2 will be the source for the next cycle.
Illustrations of outcrossing and sibmating in Cotton



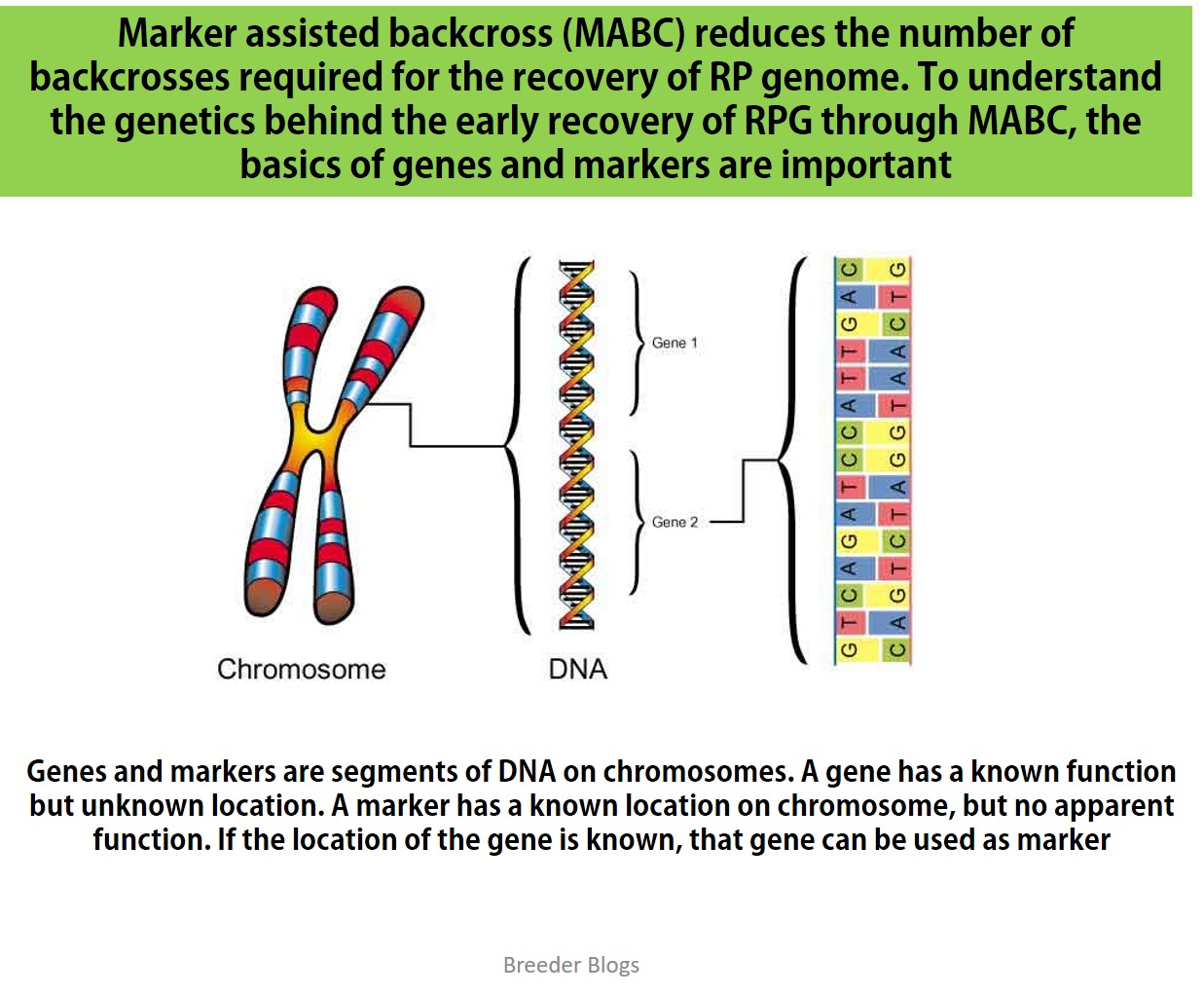














{kind=link}